
Получение рекомбинантных РНК
Получение рекомбинантных РНК обычно осуществляют методами ферментативного или химического лигирования РНК. Кроме того, недавно появилась принципиально новая возможность встраивания сегмента РНК в заданное положение других молекул РНК с помощью рибозимов.
Ковалентное сшивание отдельных сегментов РНК при получении рекомбинантных молекул, как правило, осуществляют с помощью Т4 РНК-лигазы. Т4 РНК-лигаза закодирована в геноме бактериофага Т4. Ее выделяют из клеток E.coli, зараженных этим фагом. Фермент сшивает друг с другом однотяжевые олиго- и полирибонуклеотиды. Для работы Т4 РНК-лигазы необходим источник энергии - аденозинтрифосфат. На рис. 2 приведена схема ферментативного лигирования двух коротких олигонуклеотидов. Как видно из этой схемы, акцептором в реакции лигирования служит полностью дефосфорилированный, а донором - полностью фосфорилированный по концевым нуклеотидным остаткам олигонуклеотид. Это предотвращает возможность сшивания однотипных олигонуклеотидов.
Эффективность ферментативного лигирования достаточно длинных полирибонуклеотидов сильно варьирует и ее трудно предсказать исходя только из нуклеотидной последовательности сегментов РНК. Наилучшие результаты получены в тех случаях, когда сшиваемые концы полирибонуклеотидов были пространственно сближены за счет комплементарного связывания соседних с ними участков РНК.
Недавно было установлено, что протяженные сегменты РНК (длиной в 200-300 остатков) могут быть с высоким выходом сшиты Т4 ДНК-лигазой. При этом "стыковка" сегментов осуществляется с помощью олигодезоксирибонуклеотида, комплементарного 3'-концу одного сегмента и 5'-концу другого.
Метод химического лигирования основан на активации концевой фосфатной группы одного из двух сшиваемых сегментов РНК водорастворимым карбодиимидом или
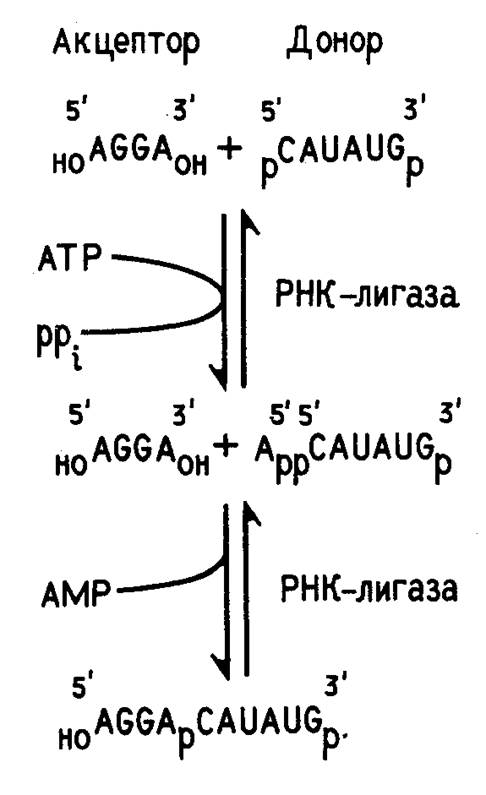
Рисунок 2. Схема сшивания двух олигорибонуклеотидов с помощью Т4 РНК-лигазы.
BrCN. В случае BrCN реакция протекает очень быстро и не сопровождается модификацией нуклеотидных остатков, хотя под действием карбодиимидов фосфодиэфирная связь образуется с более высоким выходом. Для того, чтобы обеспечить сближенность сшиваемых концевых нуклеотидных остатков в фрагментах РНК, было предложено использовать олигодезоксирибонуклеотиды, комплементарные обоим фрагментам в месте их стыка.
Химическое лигирование РНК, как правило, проходит с существенно меньшим выходом, чем ферментативное. Однако оно позволяет получать рекомбинантные РНК с необычными типами межнуклеотидной связи (например, пирофосфатной) и необычными нуклеотидными остатками в месте стыка двух фрагментов.
Получение рекомбинантных РНК с помощью рибозимов основано на обратимости реакции самосплайсинга (при отсутствии гуанозина или гуаниловых нуклеотидов). Это предоставляет возможность для встраивания интронной РНК в заданный участок другого сегмента РНК (рис. 3). Фрагмент РНК, в который производится встраивание, должен содержать нуклеотидную последовательность, идентичную нуклеотидной последовательности 3'-концевого участка 5'-экзонного района 26S РНК и соответственно комплементарную той нуклеотидной последовательности в интроне, которая отвечает за специфичность прямой реакции. Фрагмент, в который производится встраивание, берется в избытке.
В настоящее время описанная здесь цепь реакций может быть реализована только для интронной РНК, получаемой из предшественника 26S РНК тетрахимены. Однако можно думать, что конструирование новых рибозимов может существенно расширить возможности этого подхода.
